

| ����ҩ�������Ƽ��Ķ����Ŵ�������MicroRNA���ض����֬����ϳɹؼ�ø�о���չ | |||||
|
|||||
|
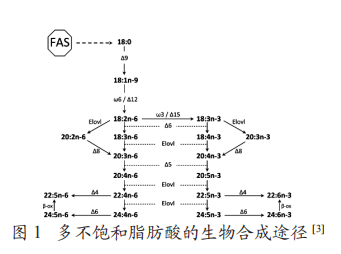
ԭ�� ��쭣������� ��ҵ��ѧ ���� ժҪ microRNA��miRNA��������������Ȼ���ڵ�һ�ַDZ���СRNA���ܹ���������ڶ����ı�������������������ĵ��ڡ����������ڳ��������֬���ᣨLC-PUFA��������ϳɣ��绨����ϩ�ᣨARA;20��4n-6������ʮ̼��ϩ�ᣨEPA;20��5n-3���Ͷ�ʮ��̼��ϩ�ᣨDHA;22��6n-3����֬����ϳ���Ҫ֬������ȥ����ø��Fads���Ĵ����������о��������й��ط����������������������֣�����ԭ�����й��ط����ּ����֬���к��д���������֬����ȷ�ζ���ʡ�Ȼ��LC-PUFA�о���һ�����չ��������Ҫ���������ཡ�����棬������������LCPUFA����ϳɹؼ�øת¼��ˮƽ���ػ��ƻ�û���γ�һ�������Ĺ۵㡣miRNA���ܲ���ת¼��ˮƽ��֬�ʴ�л���ء�Ȼ���������Ƿ�Ӱ����LC-PUFA����ϳ�;���е������в����������������һЩ����ؼ�ȥ����ø���ӳ�ø��miRNA�ڵ��ض���LC-PUFA�ϳɵı���ģʽ�͵��ػ��ơ� �ؼ��� miRNA�������֬���֬����ȥ����ø��֬�����ӳ�ø �����֬���ᣨployunsatura tedfatty acids,PUFA���ǻ��岻��ȱ�ٵı���֬���ᣬ��������≥C20�������֬���ᣨLC-PUFA������PUFA�ĸ߶����������ʽ��������ѧ�����Լ�����彡���Ĺ�ϵһֱ�ǹ��ʹ�ע���о����ȵ㡣�����о�������PUFA�������˵��n-3LC-PUFA�����ʮ̼��ϩ�ᣨEPA��20��5n-3���Ͷ�ʮ��̼��ϩ�ᣨDHA��22��6n-3�����������źŴ��ݺ͵���֬�ʴ�л����֢��Ӧ��ϸ�������Լ���ػ������ȷ��淢����Ҫ���ã�����Ԥ����������Ӳ�����γɺͷ�չ��Ԥ����Ѫ˨������Ѫ����Ѫѹ����Ѫ�ܼ���������Ϊ“Ѫ�������”������֬����͵�������֬������Ա���������������ϳɣ���PUFAͨ�������ڶ������ʳ�л�ã��������ǿ�����ijЩ������ת��Ϊ������C20-24��PUFA��LC-PUFA����C18PUFA��������ǰ��֬����ȥ����ø��Fads����֬�����ӳ�ø��ElovL����;������LC-PUFA����ͼ1������;��������͵IJ���DHA��22��5n-6�ĵ���ֱ�Ϊ18��3n-3��18��2n-6�����漰��ø��n-3��n-6ϵ��֬����������ã�һ�㶼ƫ��n-3����n-3����EPA��������ҪΔ6ȥ����ø��Δ5ȥ����ø�ֱ��α-�����ᣨALA;18��3n-3��ȥ��������18��4n-3����20��4n-3ȥ��������EPA����һ��;����ALA�ӳ�����20��3n-3�������ͨ��Δ8��Δ5ȥ����øȥ������������EPA����ijЩ���࣬����ֱ��ͨ��EPA�������22��5n-3����Δ4ȥ����ø����DHA�����ڲ��鶯���У���EPA��DHA�ƺ��漰��һ�������ӵ�·������;���Ǽ����������ձ��DHA����ϳ�;��——“Sprecherͨ·”��EPA��Ҫ��������̼���ӳ�����24��5n-3��Ȼ��ͨ����6-����ø��ȥ������������24��6n-3�����ͨ����������ø��֬�����β-��������̼�����ɶ�ʮ��̼��DHA����һ����“Δ4ȥ����øͨ·”����4;��һ�㷢���ڵ͵������У���EPAת��ΪDHA����Ҫ;����Ȼ��HuiGyuPark������ϸ����֤ʵ����ʮ��̼��ϩ�ᣨDPA��22��5n-3�����������ᣨADA��22��4n-6��ֱ��ͨ��FADS2�������ͨ����4�����γɲ����ͼ����ֱ�����DHA����Ϊ̽��FADS2�������ṩһ����˼·��Ȼ������;����������Ҫ������øA�Ȼ�ø��Acetyl CoA carboxylase,ACC����֬����ϳ�ø��fatty acid synthase,FAS�����ְ���ø���кϳɣ�Ȼ����֬������ȥ����ø��Fads���ͳ�����֬���ᣨElovL����������ø��Эͬ�����¶�֬�����һ���ӳ���ȥ���ͣ��Ӷ����̼���ϳ��IJ�����֬���ᡣ
MicroRNA��miRNA����һ����Դ�ġ��������ص�С�DZ���RNA���䳤��ԼΪ22�������ᣨnt�������е��͵ķ��л��ṹ�����������Ļ������ϣ����RNA�ۺ�øII�ӻ����������ص��������ʱ����DZ��룩�б���ij���ת¼��ת¼Ϊ����miRNA��pri-miRNA����Ȼ���ѽ��������øDrosha����ϸ�������γ�ǰ��miRNA��pre-miRNAs��Լ70nt����Ȼ��ǰmiRNA����һ������øDicer��������ϸ�����в���22nt���ȵľ��й��ܵ�miRNA�����塣�볤���DZ���RNA����״�DZ���RNA��piRNA�������DZ���RNA��ȣ�miRNA�ĵ��ػ�����Լ�ͨ���������miRNAͨ����“������”��miRNA��5'�˵�2��8���������У��ܹ���л����3'�Ƿ�������3'UTR�����ֻ���ȫ������ϣ�ʹmRNA���ȶ��������ӳٻ�����Ŀ�����ı���ڲ��鶯�������У��ݹ���ֻ��1%��5���Ļ�����ת¼������miRNA�����Ƕ��60���Ļ���ֱ�ӻ�����miRNA���ء�Ŀǰ�ڶ����Ϸ��ֺ�ȷ�ϵ�miRNA�ڲ������ࡣ�������һ��miRNA���Ե���������mRNA�ı����һ��mRNA�ı����ֿ���ͬʱ��������miRNA���ڡ����仰˵��miRNA����ͨ�����������й������ӵĵ���ϵͳ��ƾ��Դ����л���ĵ��أ�miRNA��������ڵĶ��������������ϸ������ֳ���ֻ���������Ӧ����Ӧ�����ڴ�л���źŴ�����ͬʱԽ��Խ����о�������miRNA��֬�ʴ�л��������Ҫ�ĵ������á����磬�ڲ��鶯������д��������miR-122����������ͨ�����������Ӱ�����ת�˵���1��Cpt1α���ĵ��������ڸ��൰���ʵĴ�л��ͬʱ������ͨ�����Ʋ��뵨�̴�����ϳɵĻ���ı��������ڸ���֬����͵��̴��ĺϳɡ����⣬miR-33a/b�������֬�����������ڵĹؼ�ø���������O-����ת��ø����������ת��ø1A��������-CoA-����ø��Sirtuin6��SIRT6����AMP��ø�ǻ�-α����Эͬ���ڵ��̴���̬��miR-33a/bҲ�����ȵ����������2�����Ǹ������ȵ����ź�ͨ·����Ҫ��ɲ��֣�miR-27a/bͨ������PPARγ����֬��ϸ���ֻ������������Ļ��ۣ���������е�֬����֯ʧ��������֬�ʴ�л��ػ���ת¼���Ĭ���ص���һ��ҪԪ����Ҫ����miRNA��
���������ﻹ������ţ�ȼ������ӵ�б���LC-PUFA����ϳ�;�����漰��ȥ����ø���ӳ�ø�Ļ�������ȷ�еĹ��ܺ��ܵ��DZ���RNA���ع������д�������
1 miRNA��PUFA�ϳ�;���ؼ�ȥ����ø�ĵ�������� ����ѪҺ����֯PUFAŨ������ʳ�������Դ�Ժϳɹ�ͬ������ͨ����ʳǰ�����ͣ�18:2n-6,LA����a-�����ᣨ18:3n-3,ALA���������쳤��ȥ���͡�Δ5ȥ����ø��Δ6ȥ����ø��������һø���̣��ֱ�������ͬԴֱϵ����FADS1��FADS2������롣FADS1��FADS2������FADS3����λ��Ⱦɫ��11q12-q13.1�ϵ�һ�����С��û������92kb��ɣ�ͷ��ͷ����ΪFADS1��FADS2��β��β����ΪFADS2��FADS3��FADS1��FADS2���ں���1��һ��11.4kb�����������FADS3λ��FADS2��6.0kb�����ࣨ��ͼ2����FADS3������ѧ�����в������
�����Dz���PUFA����ϳɵ���Ҫ���٣�Ҳ��FADS1��FADS2�������Ҫ��λ��Li�����о�20�����30����ĸ���ĸ���microRNA��������У��״η���FADS1��miR-365-3p��miR-218-5p��miR-181a-5p��miR-181b-5p��miR-29a-3p��miR-23b-3p����FADS2��miR-30c-1-3p���������о�ΪmiRNA��֬�ʴ�л���ӵ���ϵͳ�е���ϸ�����ṩ�˻�����Դ��Zhang��ͨ��˫ӫ����ø����֤���ˣ�mir-17�ں���ԭ����ϸ����Schl����ֱ�Ӱб�Δ4FAD��3'UTR��Ӱ����FAD1��FADS2�ı��������LC-PUFA����ϳɵĵ��ء�����ֱ�Ӱ������⣬���м�ӵ���FADS���λ�����������miR-33��Schl�а����ȵ����յ��Ļ���1��insig1���������LC-PUFA����ϳɵļ�������������Ԫ����ϵ���-1��Sembp1���ĵ���ˮ�⼤��Ӷ�����LC-PUFA����ϳɵĹؼ�øΔ4��Δ6��Δ5֬��ȥ����ø��FADs���ı������֮�⣬Chen��Ҳ����ͨ��Insig1/Srebp1ͨ·�����LC-PUFA����ϳɵĻ���miR-24��
�����ȼ������У���������̥ʱ�ڣ����dz������꣬ÿ������ʱ�ڵĸδ�л����ת����һ�����ӵķ��ӵ���ϵͳ�����磬�շ�����С���ڷ�������Щ�ѻ�֬�ʻ�Ѹ�����Ĵ������³´�лת��Ϊ��̼ˮ������Ϊ������������Դ����������ϵͳ������������������꼦������Ч���������ϣ�ͨ�������ӳ�ι��48h����ֹ�����лת���������ӳ�ι������miR-20b�������ӣ�����б�FADS1�ı����෴�ؼ��٣�������ת¼ǰ��FADS1�����ܵ�miRNA�鵼����������һת����ͬʱmiR-33�ı����������ͣ�miR-33�Ǽ�������������֬�ʴ�л���ڼ������б����ڼ��б�֤���������Ƹ���֬�������������ã��˽����һ��֧����miR-33�ڵ��ڸ���֬�ʴ�л�е����ã��������Ǽ���л���ص���Ҫת¼��������ӡ���ʾ�ڸδ�лת��ʱ�ڣ�֬����Ĵ�ͷ�ϳɿ��Լ��������ƣ����������SREBF1��������FADS�ı���������£���miRNAs�����ִ�лת���������Ǹ����лͨ·�Ĺؼ��������ӡ�
2 miRNA��PUFA�ϳ�;���ؼ��ӳ�ø�ĵ�������� ֬�����ӳ�ø��elongase of very long chain fattyacids,ELOVL����λ���������ϵ�Ĥ��ϵ���ø����Ҫ�����ӳ�ѭ����Ӧ�еĵ�һ�����Ϸ�Ӧ�����鶯���з��ֵ��ӳ�ø����������࣬����7����Ա���ֱ�ΪELOVL1~ELOVL7����Բ�ͬ��֬���������ֳ���ͬ�ĵ��������ԡ�ELOVL1��ELOVL3��ELOVL6��ELOVL7��Ҫ�ӳ������Լ���������֬���ᣬ�������볤��PUFA���쳤����ELOVL2��ELOVL4��ELOVL5������ȫ��cDNA���о��и߶�ͬԴ�ԣ�����Ҳ���������ԣ���Ҫ�������֬������ӳ���Ӧ��C18��C20PUFA�����̼�����졣
2.1 ELOVL2 ELOVL2�dz��������֬����ϳ��еĹؼ��ӳ�ø������Ҫ����DHAǰ�����ϳɹ��̣�����EPAת��Ϊ��ʮ��̼��ϩ�ᣨDPA,22��5n-3�����Լ���һ���ӳ�Ϊ24��5n-3��Ȼ����С���ELOVL2�ܹ���һ���̶��Ͽ��ӳ�18��3n-6���������ELOVL2���ܣ��������ͬ����֮����ܴ���һЩ���ܲ��졣����ԭ���Ը�ϸ����PHHs��ȥ�ֻ����о��У�������ELOVL2��ȥ�ֻ�PHHs���µ������볬����PUFAŨ�Ƚ���һ�£����⣬��������ȥ�ֻ������г��ֶ���miRNA��������ʾ��PHHsȥ�ֻ�������miRNAͨ������ELOVL�������PHH֬�ʵĺϳɡ����ۺͷ��ڡ�
2.2 ELOVL5 ELOVL2��ELOVL5ø�ڲ��鶯��Ĵ������֯���ձ�������ELOVL5�ڴ����п��ܴ������ֲ��죬�ڷκʹ����б�����ߣ��������ELOVL5��غ����������б�������ߣ�������Ϊ22��5n-6ˮƽ��Խϸߡ��������ӳ�ø�����ص����������ܣ�ELOVL5�ܹ��ӳ�C18C20PUFA������C22û�л��ԣ���ELOVL2�ܹ��ӳ�C20��C22PUFA����ELOVL2��ELOVL5�Ա���֬����͵�������֬���ᶼû�л��ԡ�
�����о�֤��ELOVL5��C18��C20Ϊ�����֬�����ת�����Ծ�����Ҫ�ԡ�Chen���״η���miR-146a����ͨ������ELOVL5�������SCHLϸ��LC-PUFA����ϳɣ�������miR-146a����������������������ELOVL5��PUFA�ӳ�ָ������20��3n-6/18��3n-6��20��4n-3/18��4n-3��22��5n-3/20��5n-3���Ӷ�������SCHLϸ����LC-PUFA������
�������±���ELOVL5ͨ���������֬��ϸ����������֬��ø��Ҳ��Ϊ��������֬ø�ṹ��patatin-like phospholipase domain���ķ�ȣ��Ӷ����ڸ���������TG��ˮƽ������֬�ʴ�л�����ڴ�֬�ʴ�л���ڹ�����miRNA���ܷ���������������á�Li��ͨ��������Ϣѧ����Ԥ���ELOVL5��miR-21-3p�İл������ʵ���б�֤ʵ����������Ƥϸ����MECs���б���bta-miR-21-3p�ٽ��˸��������IJ������������л���ELOVL5�ĵ����йء�����֮�⣬Zhang���ڴ��Լ��ش����IJ����ڼ���ԭ���Ը�ϸ���з���6��miRNA�����µ�������֤miR-218-5p����ֱ�Ӱ���ELOVL5�����ִƼ��ؿ����ϵ�ELOVL5�ı����Ӵٽ�����ĸ��������LCPUFA�ĺϳɣ����˵��ؿ���������miRNAs������������ELOVL5�ɱ����miRNAֱ�ӵ���Ӱ��LCPUFA�ĺϳɣ�ͬʱELOVL5Ҳ�����ܵ��������ӱ�������miRNA�鵼��Ӱ�죬��Ҳ���˵����ELOVL5��֬����̼ˮ������Ĵ�л����Ҫ�ԡ�
2.3 ELOVL6 ELOVL6����Ҫ�����Ǵ���������֬����ĺϳɣ������C16֬��������ΪC18������Ҫ�ڸ��ࡢ֬����֯�������ٵ�֬�ʺ����ϸߵ���֯�������б���ϸߡ�ELOVL6������Ӱ��֬����ϳɵĹؼ������ǽ������ת���ɽ�һ�������Ӳ֬�������ø������֮�⣬ELOVL6���Dz���ά�ֻ���֬�����лƽ��Ĺؼ����ӣ������µ��о��з���ELOVL6���ȵ��صֿ����Ǿƾ���֬���κ���������Ӳ���ȷ��������й����������о�ELOVL6�����������˽������֬�������ɱ仯���ơ�
���б���ELOVL6�ڹ�������ø����ֳ�D�����壨PPAR����֬�����л�ź�ͨ·�з��ӹؼ����á�ELOVL6��������ֱ�ӵ��ػ���Ĵ�л���ã���������������������з������á���HepG2ϸ����̼ˮ�����ﷴӦԪ����ϵ��ף�carbohydrate response element binding protein,ChREBP���̴�����Ԫ����ϵ���-1C��sterol regulatory elementbinding protein-1C,SREBP-1C������λ���������Эͬ�̼�ELOVL6�����ӣ�Moon��[33]���о�����ELOVL6�ı��ﻹ����ͬʱ�ܵ�SREBP-1��X����α��LXRα����ֱ�ӵ��أ���SREBP-1a��ELOVL6���յ����ø����������⣬ELOVL6�ڲ�ͬ������֯�л����Ա�miR-22-3P��mir-144��miRNA-125a-5p�ȶ���miRNAֱ�ӵ��أ���ELOVL6���ﱻ����ʱ������ᣨC16��0�������������ӣ������ᣨC18��1��n-9������ʮ̼ϩ�ᣨC20��1��n-9���Ͷ�ʮ̼��ϩ�ᣨC20��3�������������ͣ�����miRNAs����Ӱ��ELOVL6�ı���������һЩϸ���б���֬����ͳ���������֬����ĺ�����������ˣ���֬��ϸ�������������о��У�֤ʵ��ELOVL6ΪmiR�\204��ֱ�Ӱл��������֬��ϸ���ij�֬�ֻ����²�ELOVL6������mir-204��֬���ֻ�ֱ�ӵ�������֮����м�������ӡ�
3 miRNA��Ӳ֬����øAȥ����ø��SCD���ĵ�������� ��������֬�����Ǻϳ�PUFA�ĵ��Ӳ֬����øAȥ����ø��Stearoyl-CoA desaturease,SCD��Ҳ��ΪΔ9ȥ����ø���Ǹ�ϸ��������֬���ᣨSaturated fatty acid,SFA��C9�����γ�n-9ϵ��������֬���ᣨmonounsaturated fatty acid, MUFA���Ĺؼ�����ø���ֱ�ת���������ᣨOA��C18��1����������ᣨPA��C16��1���������֬���ᣨPUFA��������������������Ҫ��Դ������������Ĥ����Ҫ��ɳɷ֡��������������Ǻϳ�Ĥ��֬������֬���������������͵��̴����ȣ�����Ҫ�����SCD����ı������ø���Եĸı佫��Ӱ��ϸ���ڱ���֬�����벻����֬����ı���������Ӱ��Ĥ��֬����ɣ����ά��ϸ��Ĥ�������Լ��źŴ��ݵȺ���Ҫ��
������ָ��SCD����仯��Ӳ֬�������ı����ı�ֱ����أ��⽫��Ӱ������Ĥ���������Լ��ź�ת�����̣���һ��Ӱ��ϸ����������ֻ��������ѱ����Ĺ��������ᡢ�Ҵ�����̴����صȶ�����Դ���ػ�Ӱ�����SCD�Ļ����⣬������Դ����Ҳ��ı�SCD�Ĺ��ܱ��Tan���о�����miR-199a-3p����ֱ�Ӱ������Ӳ֬����øaȥ����ø��SCD���ı���Ӷ�������3T3-L1֬��ϸ����֬�ʻ��ۣ�������֬��ת¼��֬����ϳɣ�����Ӱ��֬����ת�˵�֬��ϸ�������Ի���ı��������½�����֬�����������̵õ�����ǿ��SCD��ֱ�Ӱ�����ػ���miR-125b��miR-192��miR-29a��������֬�λ��ۺ�MUFA����ɡ�����֮�⣬SCD�����Ժ�֬����ϳ�ø��FAS����ͬ��miR-212-5p��miR-27aֱ�Ӱ���Ӱ���ϸ������������TG���Ļ��ۡ�
����ѡ�ԡ���ҵ��ѧ��2020���6��“�Ŵ�����”��Ŀ��P106-110����Ȩ�顶��ҵ��ѧ�����У���ת�أ���ע�����������Ķ��������ݿɼ���վ��http://www.csis.cn��
����ҵ��ѧ������
�����кõ�����ϣ������ͬ�ʷ������뽫����������䣺
swineonline@263.net
��Ƹ����
����ҵ��ѧ�����¿�����־��2006������������ܵ�ҵ���ձ��ע�ͻ�ӭ�����й�֪��ͳ�ƣ��ÿ����Ϊ��ҵ�������¡�����������濯�ǡ�������֮�Ŀǰʵʩȫý��ݷñ�������ƽ̨�������ַ�չս�ԣ���Ҫ������־���꣬���������Ƹʵϰ�����ҵ����
��ϵ�ˣ������ �绰��13701057839 E_mail��delin518@x263.net
|
|||||



|